Hibiscus trionum
Hibiscus trionum
1. The products in our compound library are selected from thousands of unique natural products; 2. It has the characteristics of diverse structure, diverse sources and wide coverage of activities; 3. Provide information on the activity of products from major journals, patents and research reports around the world, providing theoretical direction and research basis for further research and screening; 4. Free combination according to the type, source, target and disease of natural product; 5. The compound powder is placed in a covered tube and then discharged into a 10 x 10 cryostat; 6. Transport in ice pack or dry ice pack. Please store it at -20 °C as soon as possible after receiving the product, and use it as soon as possible after opening.
Natural products/compounds from Hibiscus trionum
- Cat.No. Product Name CAS Number COA
-
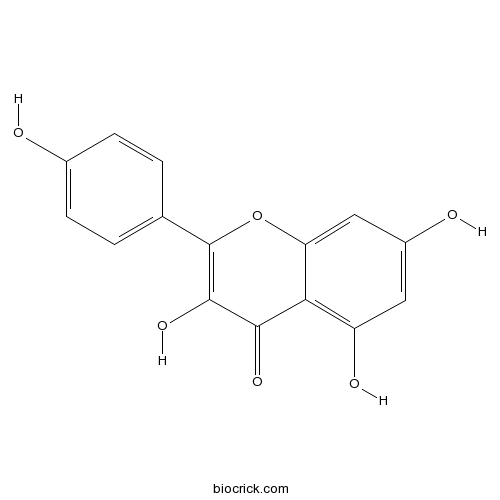
BCN5653
Kaempferol520-18-3
Instructions
-
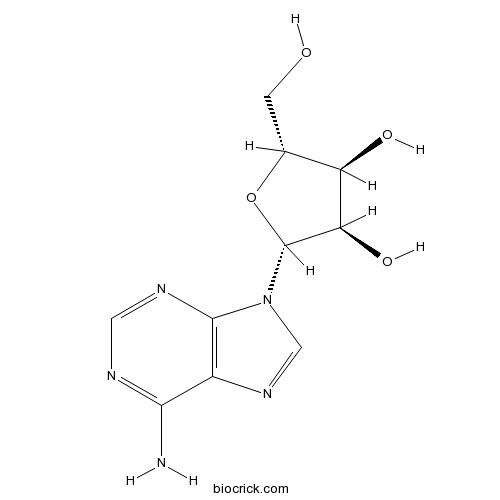
BCN5796
Adenosine58-61-7
Instructions
Cells, walls, and endless forms.[Pubmed: 27825067]
A key question in biology is how the endless diversity of forms found in nature evolved. Understanding the cellular basis of this diversity has been aided by advances in non-model experimental systems, quantitative image analysis tools, and modeling approaches. Recent work in plants highlights the importance of cell wall and cuticle modifications for the emergence of diverse forms and functions. For example, explosive seed dispersal in Cardamine hirsuta depends on the asymmetric localization of lignified cell wall thickenings in the fruit valve. Similarly, the iridescence of Hibiscus trionum petals relies on regular striations formed by cuticular folds. Moreover, NAC transcription factors regulate the differentiation of lignified xylem vessels but also the water-conducting cells of moss that lack a lignified secondary cell wall, pointing to the origin of vascular systems. Other novel forms are associated with modified cell growth patterns, including oriented cell expansion or division, found in the long petal spurs of Aquilegia flowers, and the Sarracenia purpurea pitcher leaf, respectively. Another good example is the regulation of dissected leaf shape in C. hirsuta via local growth repression, controlled by the REDUCED COMPLEXITY HD-ZIP class I transcription factor. These studies in non-model species often reveal as much about fundamental processes of development as they do about the evolution of form.
Direct surface analysis coupled to high-resolution mass spectrometry reveals heterogeneous composition of the cuticle of Hibiscus trionum petals.[Pubmed: 26335385]
Plant cuticle, which is the outermost layer covering the aerial parts of all plants including petals and leaves, can present a wide range of patterns that, combined with cell shape, can generate unique physical, mechanical, or optical properties. For example, arrays of regularly spaced nanoridges have been found on the dark (anthocyanin-rich) portion at the base of the petals of Hibiscus trionum. Those ridges act as a diffraction grating, producing an iridescent effect. Because the surface of the distal white region of the petals is smooth and noniridescent, a selective chemical characterization of the surface of the petals on different portions (i.e., ridged vs smooth) is needed to understand whether distinct cuticular patterns correlate with distinct chemical compositions of the cuticle. In the present study, a rapid screening method has been developed for the direct surface analysis of Hibiscus trionum petals using liquid extraction surface analysis (LESA) coupled with high-resolution mass spectrometry. The optimized method was used to characterize a wide range of plant metabolites and cuticle monomers on the upper (adaxial) surface of the petals on both the white/smooth and anthocyanic/ridged regions, and on the lower (abaxial) surface, which is entirely smooth. The main components detected on the surface of the petals are low-molecular-weight organic acids, sugars, and flavonoids. The ridged portion on the upper surface of the petal is enriched in long-chain fatty acids, which are constituents of the wax fraction of the cuticle. These compounds were not detected on the white/smooth region of the upper petal surface or on the smooth lower surface.
The flower of Hibiscus trionum is both visibly and measurably iridescent.[Pubmed: 25040014]
Living organisms can use minute structures to manipulate the reflection of light and display colours based on interference. There has been debate in recent literature over whether the diffractive optical effects produced by epoxy replicas of petals with folded cuticles persist and induce iridescence in the original flowers when the effects of petal pigment and illumination are taken into account. We explored the optical properties of the petal of Hibiscus trionum by macro-imaging, scanning and transmission electron microscopy, and visible and ultraviolet (UV) angle-resolved spectroscopy of the petal. The flower of Hibiscus trionum is visibly iridescent, and the iridescence can be captured photographically. The iridescence derives from a diffraction grating generated by folds of the cuticle. The iridescence of the petal can be quantitatively characterized by spectrometric measurements with several square-millimetres of sample area illuminated. The flower of Hibiscus trionum has the potential to interact with its pollinators (honeybees, other bees, butterflies and flies) through iridescent signals produced by its cuticular diffraction grating.
Contributions of iridescence to floral patterning.[Pubmed: 19641739]
The Hibiscus trionum flower is distinctly patterned, with white petals each with a patch of red pigment at the base, producing a 'bulls-eye' pattern on the whole flower. The red pigmented patches are also iridescent, due to the presence of a series of overlying cuticular striations that act as a diffraction grating. We have previously reported that scanning electron microscopy revealed a sharply defined difference between the surface structure overlying the pigmented patch and that over the rest of the petal, with the diffraction grating only present over the pigmented region. Here we show that differences in petal surface structure overlie differences in pigment color in three other species, in a range of different patterns. Floral patterns have previously been shown to be advantageous in pollinator attraction, and we discuss whether emphasis of pigment patterns by structural color may increase floral recognition by pollinators.
Floral iridescence, produced by diffractive optics, acts as a cue for animal pollinators.[Pubmed: 19119235]
Iridescence, the change in hue of a surface with varying observation angles, is used by insects, birds, fish, and reptiles for species recognition and mate selection. We identified iridescence in flowers of Hibiscus trionum and Tulipa species and demonstrated that iridescence is generated through diffraction gratings that might be widespread among flowering plants. Although iridescence might be expected to increase attractiveness, it might also compromise target identification because the object's appearance will vary depending on the viewer's perspective. We found that bumblebees (Bombus terrestris) learn to disentangle flower iridescence from color and correctly identify iridescent flowers despite their continuously changing appearance. This ability is retained in the absence of cues from polarized light or ultraviolet reflectance associated with diffraction gratings.
Tillage and residue burning affects weed populations and seed banks.[Pubmed: 17390813]
An integrated weed management approach requires alternative management practices to herbicide use such as tillage, crop rotations and cultural controls to reduce soil weed seed banks. The objective of this study was to examine the value of different tillage practices and stubble burning to exhaust the seed bank of common weeds from the northern grain region of Australia. Five tillage and burning treatments were incorporated in a field experiment, at Armidale (30 degrees 30'S, 151 degrees 40'E), New South Wales, Australia in July 2004 in a randomized block design replicated four times. The trial was continued and treatments repeated in July 2005 with all the mature plants from the first year being allowed to shed seed in their respective treatment plots. The treatments were (i) no tillage (NT), (ii) chisel ploughing (CP), (iii) mould board ploughing (MBP), (iv) wheat straw burning with no tillage (SBNT) and (v) wheat straw burning with chisel ploughing (SBC). Soil samples were collected before applying treatments and before the weeds flowered to establish the seed bank status of the various weeds in the soil. Wheat was sown after the tillage treatments. Burning treatments were only initiated in the second year, one month prior to tillage treatments. The major weeds present in the seed bank before initiating the trial were Polygonum aviculare, Sonchus oleraceus and Avena fatua. Tillage promoted the germination of other weeds like Hibiscus trionum, Medicago sativa, Vicia sp. and Phalaris paradoxa later in the season in 2004 and Convolvulus erubescens emerged as a new weed in 2005. The MBP treatment in 2004 reduced the weed biomass to a significantly lower level of 55 g/m2 than the other treatments of CP (118 g/m2) and NT plots (196 g/m2) (P < 0.05). However, in 2005 SBC and MBP treatments were similar in reducing the weed biomass. In 2004, the grain yield trend of wheat was significantly different between CP and NT, and MBP and NT (P < 0.05) with maximum yield of 5898 kg/ha in CP and 5731 kg/ha in MBP. Rainfall before the start of the second trial season promoted the germination of a large numbers of weeds. SBC and MBP treatments reduced the numbers of most of the individual weed species compared with CP, SBNT and NT. SBC was able to destroy a large proportion of seeds most likely through burning and burying some in the soil and was found to be the best treatment in exhausting the seed bank followed closely by MBP which probably buried large number of seeds deep in the soil and promoted others to germinate. CP might have buried some of the seeds in the top 5-10 cm but also promoted parts of the seed bank to germinate. SBNT and NT provided an ideal medium for weeds to germinate and resulted in heavy infestations of weeds.
