Dynamin inhibitory peptidePeptide inhibitor of GTPase dynamin CAS# 251634-21-6 |

- Laminin (925-933)
Catalog No.:BCC1015
CAS No.:110590-60-8
- Epidermal Growth Factor Receptor Peptide (985-996)
Catalog No.:BCC1014
CAS No.:96249-43-3
Quality Control & MSDS
3D structure
Package In Stock

Number of papers citing our products

| Cas No. | 251634-21-6 | SDF | Download SDF |
| PubChem ID | 90488708 | Appearance | Powder |
| Formula | C47H80N18O14 | M.Wt | 1121.26 |
| Type of Compound | N/A | Storage | Desiccate at -20°C |
| Synonyms | Gln-Val-Pro-Ser-Arg-Pro-Asn-Arg-Ala-Pro;glutaminyl-valyl-prolyl-seryl-arginyl-prolyl-asparagyl-arginyl-alanyl-proline;QVPSRPNRAP | ||
| Solubility | Soluble to 1 mg/ml in water | ||
| Sequence | QVPSRPNRAP | ||
| Chemical Name | (2S)-1-[(2S)-2-[[(2S)-2-[[(2S)-4-amino-2-[[(2S)-1-[(2S)-5-(diaminomethylideneamino)-2-[[(2S)-2-[[(2S)-1-[(2S)-2-[[(2S)-2,5-diamino-5-oxopentanoyl]amino]-3-methylbutanoyl]pyrrolidine-2-carbonyl]amino]-3-hydroxypropanoyl]amino]pentanoyl]pyrrolidine-2-carbonyl]amino]-4-oxobutanoyl]amino]-5-(diaminomethylideneamino)pentanoyl]amino]propanoyl]pyrrolidine-2-carboxylic acid | ||
| SMILES | CC(C)C(C(=O)N1CCCC1C(=O)NC(CO)C(=O)NC(CCCN=C(N)N)C(=O)N2CCCC2C(=O)NC(CC(=O)N)C(=O)NC(CCCN=C(N)N)C(=O)NC(C)C(=O)N3CCCC3C(=O)O)NC(=O)C(CCC(=O)N)N | ||
| Standard InChIKey | LHQOQXYEPUSHJL-LNIRUYPRSA-N | ||
| Standard InChI | InChI=1S/C47H80N18O14/c1-23(2)35(62-36(69)25(48)14-15-33(49)67)44(77)64-19-7-12-31(64)41(74)61-29(22-66)39(72)59-27(10-5-17-56-47(53)54)43(76)63-18-6-11-30(63)40(73)60-28(21-34(50)68)38(71)58-26(9-4-16-55-46(51)52)37(70)57-24(3)42(75)65-20-8-13-32(65)45(78)79/h23-32,35,66H,4-22,48H2,1-3H3,(H2,49,67)(H2,50,68)(H,57,70)(H,58,71)(H,59,72)(H,60,73)(H,61,74)(H,62,69)(H,78,79)(H4,51,52,55)(H4,53,54,56)/t24-,25-,26-,27-,28-,29-,30-,31-,32-,35-/m0/s1 | ||
| General tips | For obtaining a higher solubility , please warm the tube at 37 ℃ and shake it in the ultrasonic bath for a while.Stock solution can be stored below -20℃ for several months. We recommend that you prepare and use the solution on the same day. However, if the test schedule requires, the stock solutions can be prepared in advance, and the stock solution must be sealed and stored below -20℃. In general, the stock solution can be kept for several months. Before use, we recommend that you leave the vial at room temperature for at least an hour before opening it. |
||
| About Packaging | 1. The packaging of the product may be reversed during transportation, cause the high purity compounds to adhere to the neck or cap of the vial.Take the vail out of its packaging and shake gently until the compounds fall to the bottom of the vial. 2. For liquid products, please centrifuge at 500xg to gather the liquid to the bottom of the vial. 3. Try to avoid loss or contamination during the experiment. |
||
| Shipping Condition | Packaging according to customer requirements(5mg, 10mg, 20mg and more). Ship via FedEx, DHL, UPS, EMS or other couriers with RT, or blue ice upon request. | ||
| Description | Peptide inhibitor of the GTPase dynamin; competitively blocks binding of dynamin to amphiphysin thus prevents endocytosis when administered intracellularly. Reduces GABAA receptor internalization and increases miniature ISPC amplitude and frequency in neurons expressing GABAA receptors. Cell-permeable version also available. |

Dynamin inhibitory peptide Dilution Calculator
Dynamin inhibitory peptide Molarity Calculator

Calcutta University

University of Minnesota

University of Maryland School of Medicine

University of Illinois at Chicago

The Ohio State University

University of Zurich

Harvard University

Colorado State University

Auburn University

Yale University

Worcester Polytechnic Institute

Washington State University

Stanford University

University of Leipzig

Universidade da Beira Interior

The Institute of Cancer Research

Heidelberg University

University of Amsterdam

University of Auckland

TsingHua University

The University of Michigan

Miami University

DRURY University

Jilin University

Fudan University

Wuhan University

Sun Yat-sen University

Universite de Paris

Deemed University

Auckland University

The University of Tokyo

Korea University
Dynamin Inhibitory Peptide is a peptide (Gln-Val-Pro-Ser-Arg-Pro-Asn-Arg-Ala-Pro) inhibitor of the GTPase dynamin.
Dynamin is a 100-kDa large GTPase that functions to tabulate membranes and liberate nascent vesicles from the Golgi apparatus and plasma membrane. Dynamin also plays a role in many processes including division of organelles,cytokinesis and microbial pathogen resistance.
Dynamin inhibitory peptide competitively blocks binding of dynamin to amphiphysin thus prevents endocytosis when administered intracellularly. Reduces GABAA receptor internalization and increases miniature ISPC amplitude and frequency in neurons expressing GABAA receptors.
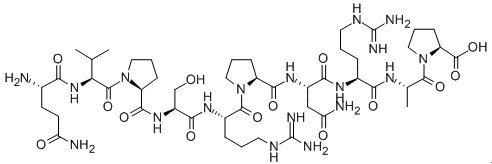
Figure1 Formula of Dynamin Inhibitory Peptide
Ref:
1. McNiven, M. A., Cao, H., Pitts, K. R. & Yoon, Y. (2000) Trends Biochem. Sci. 25, 115–120.
2. Hinshaw, J. E. (2000) Annu. Rev. Cell Dev. Biol 16, 483–519.
3. Thoms S, Erdmann R (Oct 2005). "Dynamin-related proteins and Pex11 proteins in peroxisome division and proliferation.". FEBS J 272 (20): 5169–81.
4. Grabs et al (1997) The SH3 domain of amphiphysin binds the proline-rich domain of dynamin at a single site that defines a new SH3 binding consensus sequence. J.Biol.Chem. 272 13419-25.
5. Kittler et al (2000) Constitutive endocytosis of GABAA receptors by an association with the adaptin AP2 complex modulates inhibitory synaptic currents in hippocampal neurons. J.Neurosci. 20 7972-7.
6. Nong et al (2003) Glycine binding primes NMDA receptor internalization. Nature 422 302-7.
- Isohomoarbutin
Catalog No.:BCN7612
CAS No.:25162-30-5
- Loline
Catalog No.:BCN2003
CAS No.:25161-91-5
- Acevaltrate
Catalog No.:BCN7127
CAS No.:25161-41-5
- 5-Chloro-2-nitrobenzoic acid
Catalog No.:BCC8743
CAS No.:2516-95-2
- Tesaglitazar
Catalog No.:BCC7828
CAS No.:251565-85-2
- M344
Catalog No.:BCC2162
CAS No.:251456-60-7
- SU 16f
Catalog No.:BCC7639
CAS No.:251356-45-3
- Urotensin II (human)
Catalog No.:BCC5796
CAS No.:251293-28-4
- Crebanine
Catalog No.:BCN5117
CAS No.:25127-29-1
- FRATide
Catalog No.:BCC5821
CAS No.:251087-38-4
- Antazoline HCl
Catalog No.:BCC4627
CAS No.:2508-72-7
- Salvisyrianone
Catalog No.:BCN4821
CAS No.:250691-57-7
- AM 1172
Catalog No.:BCC7675
CAS No.:251908-92-6
- SLV 320
Catalog No.:BCC7656
CAS No.:251945-92-3
- A 205804
Catalog No.:BCC3944
CAS No.:251992-66-2
- AZD7545
Catalog No.:BCC4294
CAS No.:252017-04-2
- Fmoc-D-Threoninol
Catalog No.:BCC2702
CAS No.:252049-02-8
- Fmoc-Lys(Me2)-OH
Catalog No.:BCC2567
CAS No.:252049-10-8
- Tertiapin-Q
Catalog No.:BCC5740
CAS No.:252198-49-5
- 1-Cinnamoylpyrrole
Catalog No.:BCN4006
CAS No.:252248-89-8
- 9,9'-O-Isopropyllidene-isolariciresinol
Catalog No.:BCN1474
CAS No.:252333-71-4
- Isotaxiresinol 9,9'-acetonide
Catalog No.:BCN4663
CAS No.:252333-72-5
- Lucidadiol
Catalog No.:BCN7142
CAS No.:252351-95-4
- Lucidal
Catalog No.:BCN3206
CAS No.:252351-96-5
Activity-dependent regulation of NMDA receptors in substantia nigra dopaminergic neurones.[Pubmed:24344168]
J Physiol. 2014 Feb 15;592(4):653-68.
N-Methyl-d-aspartate receptors (NMDARs) are Ca(2+)-permeable glutamate receptors that play a critical role in synaptic plasticity and promoting cell survival. However, overactive NMDARs can trigger cell death signalling pathways and have been implicated in substantia nigra pars compacta (SNc) pathology in Parkinson's disease. Calcium ion influx through NMDARs recruits Ca(2+)-dependent proteins that can regulate NMDAR activity. The surface density of NMDARs can also be regulated dynamically in response to receptor activity via Ca(2+)-independent mechanisms. We have investigated the activity-dependent regulation of NMDARs in SNc dopaminergic neurones. Repeated whole-cell agonist applications resulted in a decline in the amplitude of NMDAR currents (current run-down) that was use dependent and not readily reversible. Run-down was reduced by increasing intracellular Ca(2+) buffering or by reducing Ca(2+) influx but did not appear to be mediated by the same regulatory proteins that cause Ca(2+)-dependent run-down in hippocampal neurones. The NMDAR current run-down may be mediated in part by a Ca(2+)-independent mechanism, because intracellular dialysis with a dynamin-inhibitory peptide reduced run-down, suggesting a role for clathrin-mediated endocytosis in the regulation of the surface density of receptors. Synaptic NMDARs were also subject to current run-down during repeated low-frequency synaptic stimulation in a Ca(2+)-dependent but dynamin-independent manner. Thus, we report, for the first time, regulation of NMDARs in SNc dopaminergic neurones by changes in intracellular Ca(2+) at both synaptic and extrasynaptic sites and provide evidence for activity-dependent changes in receptor trafficking. These mechanisms may contribute to intracellular Ca(2+) homeostasis in dopaminergic neurones by limiting Ca(2+) influx through the NMDAR.
[Endocytosis of cholinoreceptors in the mechanism of depression of cholinosensitivity in Helix lucorum neurons in a cellular model of habituation].[Pubmed:20469596]
Zh Vyssh Nerv Deiat Im I P Pavlova. 2010 Mar-Apr;60(2):206-16.
Inhibitors of dynamin-dependent endocytosis (Dynamin inhibitory peptide and dynasore) and inhibitors of tubulin (colchicine and vinblastine) decrease the depression of acetylcholine-induced inward current in command neurons of Helix defensive behavior under conditions of rhythmical local applications of acetylcholine to a neuron soma in a cellular model of habituation. Mathematical model which allows for the possibility of different localizations of receptors in a neuron and characteristics of receptor travel makes it possible to analyze the dependence of the current depression on some intracellular processes. We suggest that the cholinosensitivity depression of extrasynaptic membrane zones in command neurons of Helix defensive behavior in the cellular model of habituation under study is associated with dynamin-dependent endocytosis of cholinoreceptors with involvement of cytoskeleton microtubules.
Pharmacological Investigation of Fluoro-Gold Entry into Spinal Neurons.[Pubmed:26102354]
PLoS One. 2015 Jun 23;10(6):e0131430.
The fluorescent tracer Fluoro-Gold has been widely used to label neurons retrogradely. Here we show that Fluoro-Gold can also enter neurons through AMPA receptor endocytosis. We found that a 30 minute application of Fluoro-Gold to the isolated spinal cord labeled neurons under control conditions and in the presence of glutamatergic agonists including NMDA and AMPA. The labeling was abolished or greatly reduced by glutamatergic antagonists and the endocytic inhibitors Dynasore and Dynamin inhibitory peptide. Whole cell recordings from spinal neurons exposed to extracellular AMPA revealed large inward currents that spontaneously decayed in the presence of the agonist but were maintained when a Dynamin inhibitory peptide was included in the electrode. These findings suggest that Fluoro-Gold enters spinal neurons through AMPA-mediated receptor internalization. Drugs used to induce locomotor-like activity in the spinal cord also increased and decreased Fluoro-Gold labeling in a drug and lamina specific manner, indicating that AMPAR endocytosis is altered in the presence of the locomotor cocktail. Our findings suggest that endocytosis of Fluoro-Gold could potentially complicate the interpretation of experiments in which the tracer is used to label neurons retrogradely. Moreover, they also demonstrate that many drugs, including the locomotor cocktail, can modulate the number and/or the composition of AMPA receptors on spinal neurons and thereby affect network excitability.
A late phase of LTD in cultured cerebellar Purkinje cells requires persistent dynamin-mediated endocytosis.[Pubmed:22049330]
J Neurophysiol. 2012 Jan;107(1):448-54.
Long-term synaptic depression (LTD) of cerebellar parallel fiber-Purkinje cell synapses is a form of use-dependent synaptic plasticity that may be studied in cell culture. One form of LTD is induced postsynaptically through an mGlu1/Ca influx/protein kinase Calpha (PKCalpha) cascade, and its initial expression requires phosphorylation of ser-880 in the COOH-terminal PDZ-ligand region of GluA2 and consequent binding of PICK1. This triggers postsynaptic clathrin/dynamin-mediated endocytosis of GluA2-containing surface AMPA receptors. Cerebellar LTD also has a late phase beginning 45-60 min after induction that is blocked by transcription or translation inhibitors. Here, I have sought to determine the expression mechanism of this late phase of LTD by applying various drugs and peptides after the late phase has been established. Neither bath application of mGluR1 antagonists (JNJ-16259685, LY-456236) nor the PKC inhibitor GF-109203X starting 60-70 min after LTD induction attenuated the late phase. Similarly, achieving the whole cell configuration with a second pipette loaded with the peptide PKC inhibitor PKC(19-36) starting 60 min postinduction also failed to alter the late phase. Late internal perfusion with peptides designed to disrupt PICK1-GLUA2 interaction or PICK1 dimerization failed to impact late phase LTD expression. However, late internal perfusion with two different blockers of dynamin, the drug dynasore and a Dynamin inhibitory peptide (QVPSRPNRAP), produced rapid and complete reversal of cerebellar LTD expression. These findings suggest that the protein synthesis-dependent late phase of LTD requires persistent dynamin-mediated endocytosis, but not persistent PICK1-GluA2 binding nor persistent activation of the upstream mGluR1/PKCalpha signaling cascade.
Phospholipase C-dependent Ca2+-sensing pathways leading to endocytosis and inhibition of the plasma membrane vacuolar H+-ATPase in osteoclasts.[Pubmed:20592242]
Am J Physiol Cell Physiol. 2010 Sep;299(3):C570-8.
In osteoclasts, elevation of extracellular Ca2+ is an endogenous signal that inhibits bone resorption. We recently found that an elevation of extracellular Ca2+ decreased proton extrusion through the plasma membrane vacuolar H+-ATPase (V-ATPase) rapidly. In this study we investigated mechanisms underlying this early Ca2+-sensing response, particularly in reference to the activity of the plasma membrane V-ATPase and to membrane retrieval. Whole cell clamp recordings allowed us to measure the V-ATPase currents and the cell capacitance (C(m)) simultaneously. C(m) is a measure of cell surface. Extracellular Ca2+ (2.5-40 mM) decreased C(m) and the V-ATPase current simultaneously. The decreased C(m), together with the enhanced uptake of a lipophilic dye (FM1-43), indicated that Ca2+ facilitated endocytosis. The endocytosis was blocked by dynamin inhibitors (dynasore and dynamin-inhibitory peptide), by small interfering RNA (siRNA) targeting for dynanmin-2 and also by bafilomycin A(1), a blocker of V-ATPases. The extracellular Ca2+-induced endocytosis and inhibition of the V-ATPase current were diminished by a phospholipase C inhibitor (U73122) and siRNA targeting for phospholipase C gamma2 subunit. Holding the cytosolic Ca2+ at either high (0.5-5 microM) or low levels or inhibiting calmodulin by an inhibitor (W7) or an antibody (anti-CaM) decreased the stimulated endocytosis and the inhibition of the V-ATPase current. These data suggest that extracellular Ca2+ facilitated dynamin- and V-ATPase-dependent endocytosis in association with an inhibition of the plasma membrane V-ATPase. Phospholipase C, cytosolic Ca2+, and calmodulin were involved in the signaling pathways. Membrane retrieval and the plasma membrane V-ATPase activity may cooperate during the early phase of Ca2+-sensing response in osteoclasts.
Glycine binding primes NMDA receptor internalization.[Pubmed:12646920]
Nature. 2003 Mar 20;422(6929):302-7.
NMDA (N-methyl-d-aspartate) receptors (NMDARs) are a principal subtype of excitatory ligand-gated ion channel with prominent roles in physiological and disease processes in the central nervous system. Recognition that glycine potentiates NMDAR-mediated currents as well as being a requisite co-agonist of the NMDAR subtype of 'glutamate' receptor profoundly changed our understanding of chemical synaptic communication in the central nervous system. The binding of both glycine and glutamate is necessary to cause opening of the NMDAR conductance pore. Although binding of either agonist alone is insufficient to cause current flow through the channel, we report here that stimulation of the glycine site initiates signalling through the NMDAR complex, priming the receptors for clathrin-dependent endocytosis. Glycine binding alone does not cause the receptor to be endocytosed; this requires both glycine and glutamate site activation of NMDARs. The priming effect of glycine is mimicked by the NMDAR glycine site agonist d-serine, and is blocked by competitive glycine site antagonists. Synaptic as well as extrasynaptic NMDARs are primed for internalization by glycine site stimulation. Our results demonstrate transmembrane signal transduction through activating the glycine site of NMDARs, and elucidate a model for modulating cell-cell communication in the central nervous system.
Constitutive endocytosis of GABAA receptors by an association with the adaptin AP2 complex modulates inhibitory synaptic currents in hippocampal neurons.[Pubmed:11050117]
J Neurosci. 2000 Nov 1;20(21):7972-7.
Type A GABA receptors (GABA(A)) mediate the majority of fast synaptic inhibition in the brain and are believed to be predominantly composed of alpha, beta, and gamma subunits. Although changes in cell surface GABA(A) receptor number have been postulated to be of importance in modulating inhibitory synaptic transmission, little is currently known on the mechanism used by neurons to modify surface receptor levels at inhibitory synapses. To address this issue, we have studied the cell surface expression and maintenance of GABA(A) receptors. Here we show that constitutive internalization of GABA(A) receptors in hippocampal neurons and recombinant receptors expressed in A293 cells is mediated by clathrin-dependent endocytosis. Furthermore, we identify an interaction between the GABA(A) receptor beta and gamma subunits with the adaptin complex AP2, which is critical for the recruitment of integral membrane proteins into clathrin-coated pits. GABA(A) receptors also colocalize with AP2 in cultured hippocampal neurons. Finally, blocking clathrin-dependant endocytosis with a peptide that disrupts the association between amphiphysin and dynamin causes a large sustained increase in the amplitude of miniature IPSCs in cultured hippocampal neurons. These results suggest that GABA(A) receptors cycle between the synaptic membrane and intracellular sites, and their association with AP2 followed by recruitment into clathrin-coated pits represents an important mechanism in the postsynaptic modulation of inhibitory synaptic transmission.
The SH3 domain of amphiphysin binds the proline-rich domain of dynamin at a single site that defines a new SH3 binding consensus sequence.[Pubmed:9148966]
J Biol Chem. 1997 May 16;272(20):13419-25.
Amphiphysin is an SH3 domain-containing neuronal protein that is highly concentrated in nerve terminals where it interacts via its SH3 domain with dynamin I, a GTPase implicated in synaptic vesicle endocytosis. We show here that the SH3 domain of amphiphysin, but not a mutant SH3 domain, bound with high affinity to a single site in the long proline-rich region of human dynamin I, that this site was distinct from the binding sites for other SH3 domains, and that the mutation of two adjacent amino acids in dynamin I was sufficient to abolish binding. The dynamin I sequence critically required for amphiphysin binding (PSRPNR) fits in the novel SH3 binding consensus identified for the SH3 domain of amphiphysin via a combinatorial peptide library approach: PXRPXR(H)R(H). Our data demonstrate that the long proline-rich stretch present in dynamin I contained multiple SH3 domain binding sites that recognize interacting proteins with high specificity.
